Researchers led by Prof. ZHANG Zai-Rong from the Interdisciplinary Research Center on Biology and Chemistry, Shanghai Institute of Organic Chemistry have identified a post-translational topogenesis pathway for the folding and assembly of multi-spanning membrane proteins (MSPs) (Fig. 1).Of the approximately 5000 membrane proteins synthesized at the ER membrane of human cells, over half are MSPs that play crucial roles in cellular and organismal physiology, functioning as ion channels, transporters, receptors, and intramembrane enzymes. A significant proportion of these functions rely on polar and charged amino acids, which lead to the formation of poorly hydrophobic TMDs (pTMDs). However, pTMDs face challenges in being recognized and integrated into the phospholipid bilayer by Sec61 translocon, which prefer hydrophobic TMDs. In the human proteome, approximately 30% of membrane proteins and more than 50% of MSPs contain at least one pTMD. How these pTMDs are effectively identified and precisely packaged into mature MSP structures has been a significant scientific question. Using the six-spanning protein adenosine triphosphate-binding cassette transporter G2 (ABCG2) as a model, researchers found that during co-translational translocation, ABCG2’s pTMD2 passes through the central pore of the translocon into the ER lumen, rather than being integrated into the phospholipid bilayer through the translocon's lateral gate. This results in the insertion of downstream TMDs into the ER membrane with reverse orientations, thereby forming a unique intermediate. Following the translation of the C-terminal positively charged twin lysine residues, a near-global topological rearrangement process occurs. Affinity purification demonstrated that ATP13A1 can detect the C-terminal positive charge signal of ABCG2. The replacement of lysine residues with negatively charged or neutral amino acids results in a significant weakening of the interactions between ATP13A1 and ABCG2 mutants. Furthermore, the knockout of ATP13A1 resulted in the evident accumulation of misfolded ABCG2 conformations, primarily those with misoriented TMD6 within the ER membrane. Therefore, ATP13A1 plays a crucial role in the topogenesis of MSPs, where its ATPase activity promotes the dislocation of the misoriented TMD6 from the lipid bilayer into the cytosol. Thereafter, the cytosol-exposed TMD6 is reintegrated into the ER membrane, thereby driving the post-translational topological rearrangement of other upstream TMDs. Upon successful rearrangement of TMDs 4-6, the intermediate can oligomerize into a quaternary structure. This process likely facilitates pTMD2 to integrate into the final structure from the aqueous ER lumen, yielding tightly enveloped pTMD2 by surrounding TMDs. In summary, the present study explains how a “difficult” pTMD is co-translationally skipped for insertion and post-translationally buried into the final correct structure at the late folding stage, thus avoiding excessive lipid exposure. Notably, due to the exposure of pTMD2 to the ER lumen during the topogenesis of ABCG2, the N441 glycosylation modification caused by the ABCG2-S441N genetic mutation can significantly block pTMD assembly at the late stage of topogenesis. As ABCG2 is a uric acid transporter, this study explains how this mutation is closely associated with human diseases such as gout and hyperuricemia. This work was published in Molecular Cell on May 8, 2024. The research was supported by the National Natural Science Foundation of China and the Shanghai Municipal Science and Technology Major Project. Paper link: https://authors.elsevier.com/a/1j3583vVUPRl%7Er 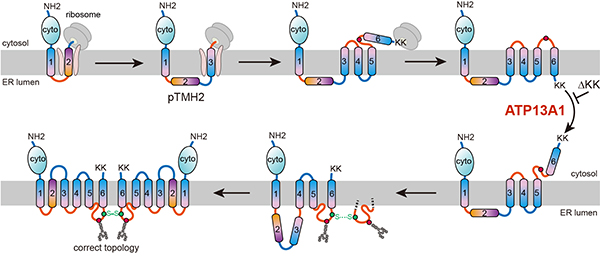
Fig. 1. Schematic representation of the ABCG2 topogenesis pathway assisted by ATP13A1. (Image by ZHANG Zai-Rong) ZHANG Zai-Rong Ph.D., Principal Investigator Interdisciplinary Research Center on Biology and Chemistry, Shanghai Institute of Organic Chemistry, Chinese Academy of Sciences Haike Road 100 Shanghai 201210 China Tel: 0086-21-68582263 Email: zrzhang@sioc.ac.cn |
